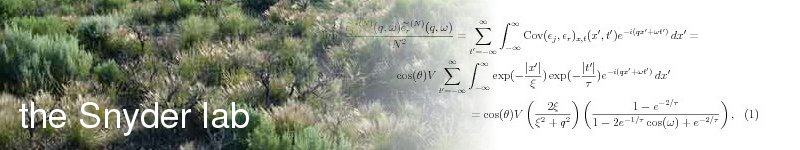 `
`
On the one hand, spatially or temporally variable environmental conditions can create new niches. By enabling species to specialize in conditions at particular places or times, environmental variation can help individuals reduce their exposure to between-species competition, sometimes allowing species to persist which would otherwise have been competitively excluded. Exploiting environmental variation in this way requires species to evolve traits that heighten their sensitivity to environmental variation. On the other hand, being sensitive to environmental variation can cause growth rates to vary in time. For organisms that reproduce in synchronized pulses, such as plants, year to year variation leads to lower long-run growth/fitness. The risks of environmental variation should lead species to evolve traits that will buffer them against variation.
Past work focused on ways that environmental variation could promote species coexistence. Current work leans toward evolutionary responses to environmental variation. For example, "Leaving home ain't easy: non-local seed dispersal is only evolutionarily stable in highly unpredictable environments" (Snyder, Proc. R. Soc. B, 2011, 278(1706), 739--744) explores the ability of environmental variation to drive the evolution of seed dispersal. Unless the environment is highly unpredictable, evolutionarily stable dispersal distances leave seeds close to their mother, in places whose environmental conditions are correlated with that of their natal location instead of in novel environments. This is an example of species increasing their sensitivity to environmental variation. "Coexistence and coevolution in fluctuating environments: Can the storage effect evolve?" (R. Snyder and P. Adler, Am. Nat., 2011, 178(4), E76--E84) considers whether two competitors can co-evolve variable germination rates and thereby benefit from a storage effect. We find that it is very difficult to coevolve variable germination if germination is not predictive (more seeds germinate in ``good'' years), though it is possible to achieve if one species has a fixed strategy, perhaps due to reduced genetic variation. Here species evolve buffering strategies unless there is some other benefit (predictive germination) to increasing their sensitivity to year-to-year variation.
Benthic marine organisms typically produce 10^many tiny larvae when they spawn. The larvae are weak swimmers. They can control their depth in the water column, enabling them to sample different currents, but beyond that, they largely go where the currents drive them. Ocean flows are highly variable and yet not without structure: eddies form over tens of kilometers and last weeks or months. That structure has implications for how variation in larval transport affects overall population growth ("How much do marine connectivity fluctuations matter?", R. Snyder, C. Paris, A. Vaz, in review).
Larvae released within a few days of each other are caught up in the same water packet and succeed or fail as a group. There are therefore fewer independent dispersing units and more demographic stochasticity than one would naively expect. This system has lead us to some unexpected conclusions about bet hedging strategies in addition to consequences for marine life history --- stay tuned! (Work in progress with W. Woyczynski and S. Burgess.)